ESTRUCTURA DEL COLÁGENO
|
|
ESTRUCTURA DEL COLÁGENO |
|
El colágeno es un material extracelular fabricado por los fibroblastos y es una proteína fibrosa que resulta relativamente insoluble en agua, en contraposición a otras familias de llamadas globulares, que sí son solubles en agua. La base molecular del colágeno está constituida por cadenas de polipéptidos y cada uno de éstos es un polímero de aminoácidos. Es decir, son cadenas constituidas por aminoácidos, que son unidades moleculares pequeñas. Cada uno de estos aminoácidos se caracterizan por tener por lo menos dos funciones distintas: una amino y una ácida en la misma unidad molecular. Los polipéptidos no son más que cadenas de estos aminoácidos que se encuentran en los organismos biológicos en números limitados. Existen unos 20 aminoácidos fundamentales. Su secuencia permite identificar las cadenas de polipéptidos. Todas las proteínas que existen en la biología se identifican porque poseen una secuencia particular de aminoácidos que es única y propia de esa proteína, que permite identificarla por métodos bioquímicos, inmunológicos, etc.
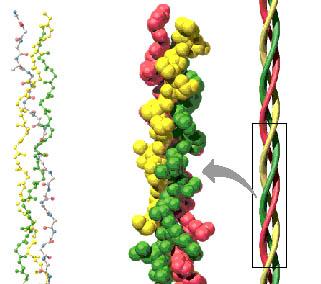
Estas macromoléculas de tropocolágeno son muy pequeñas. Sólo se conocen por métodos indirectos, son detectables bioquímicamente . Las macromoléculas de tropocolágeno se agrupan entre sí constituyendo estructuras llamadas fibrillas de colágeno. Cada fibrilla de colágeno está constituida por miles de moléculas de tropocolágeno, que son visibles al microscopio electrónico, se pueden detectar, medir, colorear, estudiar en forma relativamente cómoda. Si bien en algunas partes están aisladas, más o menos sueltas, en la mayor hurte del organismo, sobre todo en la dermis, centenares de estas fibrillas se unen lado a lado formando fibras colágenas mucho más voluminosas, visibles con microscopio óptico. Las fibras colágenas tienden a agruparse en conjuntos más grandes llamados haces colágenos. Antes de entrar en más detalles de la constitución química de esta macromolécula vamos a ver algunas propiedades físicas que son importantes. En primer lugar, el colágeno está especialmente concentrado en aquellos tejidos que soportan peso (el peso del organismo), fundamentalmente los cartílagos y los huesos. También existe colágeno concentrado en altas proporciones en aquellas partes del organismo que transmiten fuerza, como los tendones (ligamentos que unen los músculos con las piezas esqueléticas). En tercer lugar, el colágeno aparece en forma numerosa en aquellos lugares como la dermis o las fascias (láminas que recubren los músculos) sirven pura proteger, o donde se necesita un material que resista la tracción o los cambios de volumen. Finalmente, el colágeno, en una de sus formas, constituye prácticamente una armazón de microfibrillas, que sostiene la estructura de todos los órganos y vísceras del organismo. 0 sea, que encontramos pequeñas fibrillas de colágeno en el hígado, en el bazo, en el pulmón, en fin, no hay ninguna víscera que no tenga esa armazón de colágeno. Por ejemplo los vasos sanguíneos que son tubos, mantienen su forma tubular en función de que alrededor de la pared tienen una trama en espiral de fibrillas de colágeno, que constituyen el soporte de esa estructura tubular. En general, el colágeno aparece como un material altamente ordenado. En algunos lugares las fibras de colágeno se disponen en forma estrictamente paralela. El ejemplo más típico es el de los tendones. En otros lugares como la dermis, las fibras colágenas aparecen entrelazadas en todos los planos del espacio de un modo muy apretado. De modo que cuando nosotros observamos la dermis al microscopio óptico, o sea con poca resolución y sin ningún artificio que nos permita separar las fibras de colágeno. prácticamente no podemos distinguir los límites entre una fibra y otra, porque están formando una malla demasiado apretada. Por eso la dermis vista al microscopio óptico con coloraciones normales o de rutina aparece como un tejido conjuntivo casi homogéneo, donde de vez en cuando se ven células separadas. De ahí que clásicamente para estudiar el tejido conjuntivo de la dermis se hallan ideado una serle de métodos para separar las fibras. Uno de los métodos más clásicos es separar las fibras por métodos mecánicos y hacer una disociación de las mismas con agujas, de modo de poder observarlas al microscopio con más comodidad. Pero el procedimiento también clásico pero más eficiente es producir un edema, es decir, inyectar la dermis con suero fisiológico de modo de aprovechar una de las propiedades de la sustancia fundamental amorfa que es una gran capacidad de hidratarse, de embeberse con agua y por lo tanto, aumentar el volumen y al hincharse la sustancia fundamental, las fibras colágenas aparecen más separadas En estas circunstancias, lo que vemos como material colágeno, son haces que tienden a veces a ramificarse, ondulados, que tienen dimensiones variables, poro que se miden en micras o en micrómetros, o sea en unidades que, están en el orden de 10 -6, o sea que cada micra es la milésima parte del milímetro Los haces de fibras colágenas miden habitualmente entre 15 y 30 micras, aunque hay también haces más finos y más gruesos; en cada uno de estos haces, nosotros encontramos fibrillas, fibras de colágeno que se disponen paralelamente. Cuando el haz se ramifica, las fibrillas se distribuyen, pero las fibras en sí mismas no se ramifican. Es decir, lo que se puede ramificar es el haz, pero no las fibras de colágeno. Es frecuente encontrar en estos haces, en la visión del tejido conjuntivo, células, fibroblastos que están unidos, pegados a la superficie de los haces de colágeno. Esos fibroblastos o están sintetizando nuevo material colágeno. o están reponiendo material colágeno que se está desgastando, porque una de las características de esta proteína, igual que de muchas otras del organismo, es que una vez que se forman, después que pasa el tiempo son sometidas a un constante recambio, o sea, hay partes que salen y partes que se renuevan. Otro concepto importante es el de que el colágeno forma parte de un
complejo funcional que es el tejido conjuntivo.
Clásicamente, para colorear los haces de fibras colágenas. con el microscopio óptico, se han utilizado una variedad de colorantes que son útiles para ponerlas en evidencia o para distinguirlas de otras fibras, como las musculares o mismo las elásticas. Así por ejemplo existe un colorante llamado verde luz, que colorea selectivamente a las fibras colágenas. Del mismo modo, el azul de toluidina colorea más o menos selectivamente ir las fibras colágenas. Durante bastante tiempo se hablaba solamente de un tipo de fibras colágenas. Ya hace bastantes años, se comprobó que cuando se utilizaba un método de coloración basado en la impregnación de tejidos con sales metálicas (en este caso la sal metálica más utilizada ha sido el carbonato de plata) se pueden distinguir dos tipos de fibras colágenas. Un tipo de fibras que aparecen gruesas de un color rojo, y que abundan por ejemplo en la dermis, los tendones, en las cápsulas de los órganos, etc. y fibrillas o fibras de colágeno más finas, que aparecen de color negro, que durante mucho tiempo se denominaron fibras de reticulina. 0 sea, que con métodos de tinción al microscopio óptico no podemos discriminar más que esos dos tipos de material colágeno. Cuando se utilizan métodos bioquímicos, inmunológicos y también la microscopía electrónica se ha llegado a discriminar en una primera etapa 5 tipos de fibras colágenas. y más modernamente, hasta 12. Vamos a referirnos en principio, a esos primeros 5 tipos, pero antes tenemos que hacer una breve incursión por la estructura ya más fina de estas fibras. Cuando estudiamos un pequeño fragmento del haz de fibras colágenas al microscopio óptico, observamos un panorama bastante más complicado. Observamos fibrillas de un diámetro variable, pero que anda alrededor de las 100 milimicras, o sea, una unidad que a su vez es la milésima parte de la micra, por lo tanto es 10 -9 y esas fibras que tienen la longitud muy grande, a su vez aparecen subdivididas por una serie de bandas que en el microscopio electrónico aparecen coloreadas de color oscuro o de color más claro y esas bandas se repiten a lo largo de la fibra constituyendo una. periodicidad (o sea, una estructura bandeada que se repite siempre igual a sí misma). La distancia entre las bandas que se repiten dentro de la fibrilla es siempre igual, entre 60 y 70 nanómetros. Habitualmente se habla de 64 nanómetros (esto es la periodicidad del colágeno). Y precisamente cuando se colocan estas fibras llamadas de colágeno nativo (o sea, colágeno que aparece naturalmente en los tejidos) en medio ácido, un medio con ácido acético débil, se disocian, pierden la periodicidad y aparece la unidad que las constituye, que es la molécula de tropocolágeno, que tiene una longitud de 300 mm y un diámetro de 1.4 mm (o sea, que es una macromolécula que ha sido medida). Cuando se neutraliza este medio ácido con álcali, se puede observar que las fibrillas del tropocolágano vuelven a agregarse muchas de ellas lado a lado, para reconstituir la fibrillas da colágeno nativa y vuelve a aparecer la periodicidad característica de las fibras colágenas. El peso molecular del tropocolágeno ha sido estimado entre 300.000 y 325.000. Cada molécula de tropocolágeno esta constituida por tres cadenas de polipéptidos en cada una de las cuales hay alrededor de 1000 aminoácidos. La estructura de la triple hélice del tropocolágeno es fundamental y es característica de esta proteína fibrilar. Cuando existen defectos (incluso se conocen ciertos defectos genéticos) por los cuales existen déficit en algunos aminoácidos que constituyen la cadena de polipétidos del colágeno, entonces la triple hélice no se puede formar y en esos casos la molécula de tropcolágeno es defectuosa e incapaz de reconstituir la fibrilla de colágeno (o sea, no existe o no se forma el colágeno). Eso se ve en algunas enfermedades, algunas de origen hereditario y otras producidas por sustancias químicas, drogas, etc. Cuando se analiza ya la composición química de estas cadenas de polipéptidos que constituyen el colágeno, se ve que los aminoácidos que conforman el colágeno tienen una distribución bastante regular, que es lo que caracteriza a las proteínas. Encontramos una estructura que se llama repetitiva en la secuencia de aminoácidos que se simboliza de esta manera: Gli – x – y – Gli – x – y – Gli – x- y - Gli
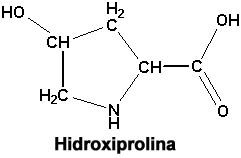
De modo que en síntesis lo que caracteriza al colágeno es esa secuencia repetitiva y la gran proporción que tiene de glicina, prolina e hidroxiprolina. La prolina y la hidroxiprolina constituyan juntas 22 % de todos los aminoácidos del colágeno. Se sabe que la hidroxiprolina desempeña un papel fundamental y especial como elemento que estabiliza esta triple hélice. Cuando hay defectos de la hidroxiprolina se traduce en la desorganización de la triple hélice y por lo tanto de todo el colágeno. Finalmente, existen otros dos aminoácidos que se encuentran solamente en el colágeno, que son lisina y la hidroxilisina. Para terminar con esta parte de la anatomía de la molécula del tropocolágeno, tenemos que hacer un pequeño agregado a esta disposición en triple hélice. Las tres moléculas están perfectamente entrelazadas a lo largo de toda la molécula de tropocolágeno menos en las puntas, aquí se pierde la triple hélice, de modo que podemos imaginar la molécula de tropocolágeno como una barra (un cilindro) y en las extremidades las tres moléculas polipéptidicas más desorganizadas y estas puntas son las que precisamente intervienen para formar uniones químicas con las moléculas de tropocolágeno adyacentes. El tropocolágeno como tal se forma en el fibroblasto y sale de él, pero la fibrilla de colágeno se forma sólo por la agregación ordenada de este tropocolágeno y esa agregación ordenada se da también de una manera muy regular y específica, que es lo que veremos ahora. Cada molécula de tropocolágeno la podemos representar de esta manera:
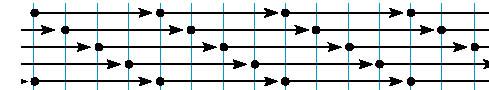
0 sea, una molécula que tiene 300 mm de longitud y que además está polarizada con dos extremidades diferentes. En un primer sentido, las moléculas de tropocolágeno se ordenan a lo largo unas de otras, pero en la segunda hilera de moléculas, en el colágeno nativo, hay una hilera que vendrá más de atrás y así sucesivamente se colocan desfasadas.
La cuarta molécula coincide con la primera. Esta disposición desfasada explica la aparición en el microscopio electrónico cuando se utilizan colorantes, una serie de bandas transversales que resultan del alineamiento en sentido transversal de distintas partes de la molécula de tropocolágeno que están dispuestas de esta manera. Esta disposición ordenada de las macromoléculas explica también una propiedad física y óptica fundamental que es la birrefingencia, que es uno de los métodos que se ha empleado para distinguir las distintas modalidades del colágeno. Revisión de los distintos tipos de colágeno:
|
<<<Regresar a Técnica del Cuero>>>
Cueronet.com
info@cueronet.com
Copyright © Cueronet.com
Reservados todos los derechos.